|
La substance grise de la moelle ťpiniŤre est divisťe en 10 couches ou lames
cytoarchitectoniques (organisation de REXED). ņ l'ťtage supťrieur, les
fibres A-delta aboutissent principalement dans la premiŤre lame et
plus rarement dans la partie superficielle de la deuxiŤme (voir figure 2.4).
Les fibres affťrentes provenant des tissus profonds et des viscŤres prennent
fin essentiellement dans les lames I et V (100). Les fibres C se terminent
principalement dans la lame I et II. Les grosses fibres myťlinisťes
A-alpha achŤvent leurs parcours dans la lame III ou dans les lames
plus profondes. En dťpit de la spťcificitť de l'entrťe des fibres A-alpha,
A-delta et C dans les lames, ces fibres ťtablissent des connexions
entre elles. Par exemple, un axone se terminant dans une lame donnťe peut
entrer en contact avec les neurones des lames voisines via une collatťrale
axonale pťnťtrant cette rťgion. ņ l'inverse, le contact s'ťtablit parfois
via le dendrite d'un neurone issu des lames voisines et s'enfonÁant dans sa
propre lame. L'axone peut aussi faire synapse avec un interneurone provenant
d'une lame voisine (27). La corne dorsale reste le site
privilťgiť d'une
importante convergence synaptique. De nombreux neurones affťrents primaires,
myťlinisťs ou non, ťtablissent des liaisons synaptiques avec quelques
neurones de la corne dorsale. Il existe trois principales catťgories
de cellules nerveuses : neurones de projection, interneurones excitateurs
et interneurones inhibiteurs.
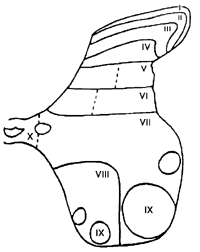
Figure 2.4 Organisation des lames
|
Trois principales catťgories de cellules nerveuses :
-
Les neurones de projection relaient le message vers l'encťphale. Ils se
divisent ŗ leur tour en neurones nociceptifs spťcifiques et en
neurones nociceptifs non-spťcifiques ŗ large gamme dynamique. Les neurones
nociceptifs spťcifiques ne reÁoivent leur information que des nocicepteurs
affťrents primaires. Leur champ rťcepteur varie entre un et quelques
centimŤtres carrťs. Les neurones nociceptifs non-spťcifiques recueillent
leur information autant des nocicepteurs affťrents primaires que des
mťcanorťcepteurs. Ils possŤdent un champ rťcepteur trŤs ťtendu qui couvre
souvent l'ensemble d'un membre. Ce champ rťcepteur se divise en trois
zones. La zone 1, centrale, ŗ partir de laquelle toute stimulation,
nociceptive ou non, provoque une excitation. En pťriphťrie de cette zone,
la zone 2 d'oý les stimulations les plus faibles dťclenchent un effet
inhibiteur et les plus intenses, un effet excitateur. Puis, la zone 3,
plus ťtendue encore, oý toute stimulation correspond ŗ un effet inhibiteur
(27). Les neurones nociceptifs spťcifiques prťvalent dans la lame I mais
se retrouvent aussi dans les lames II, IV et V. Les neurones ŗ large gamme
dynamique prťdominent dans les lames V et VI et demeurent faiblement
reprťsentťs dans les lames I, II et IV (303). On rapporte en plus la
prťsence de neurones nociceptifs ŗ projections complexes dans les lames
VII et VIII. Ils se caractťrisent par la trŤs grande convergence de
l'information qu'ils reÁoivent des deux cŰtťs du corps, incluant celle
issue des rťcepteurs viscťraux. De plus, la lame X renfermerait
principalement des neurones nociceptifs de projection qui ressembleraient
ŗ ceux des lames I et II (100).
-
Les interneurones excitateurs relaient l'information ŗ des neurones
de projection, ŗ d'autres interneurones ou ŗ des neurones moteurs
susceptibles d'induire une rťponse rťflexe spinale. Ils se concentrent
surtout au niveau de la lame II. Leur axone se termine un ou deux segments
aprŤs leur corps cellulaire. Ils font synapse localement, probablement
avec d'autres neurones de la lame II.
-
Enfin, les interneurones inhibiteurs participent, entre autres, au
contrŰle du message nociceptif et prťsentent des caractťristiques
identiques ŗ celles des interneurones excitateurs. Les lames
superficielles (I et II) contiennent une grande densitť de neurones
nociceptifs et non nociceptifs. Elles constituent ainsi une rťgion
importante pour la modulation des affťrences. Les neurotransmetteurs
interviennent aussi dans la transmission de l'influx nerveux. Plus d'une
substance jouent un rŰle dans la transmission de l'information nociceptive.
Plusieurs peptides actuellement identifiťs dans les cellules du ganglion
de la racine dorsale peuvent Ítre considťrťs comme des transmetteurs
potentiels. La substance P, la somatostatine et le polypeptide intestinal
vasoactif figurent parmi les plus probables. D'autres transmetteurs,
n'appartenant pas ŗ la famille des peptides, interviennent probablement ;
parmi eux, des acides aminťs tels le glutamate, l'aspartate ou l'adťnosine
triphosphate nuclťotide (100).
|