|
L'effet analgťsique de la stimulation des zones g‚chettes ŗ l'aide de balles
s'explique par le mťcanisme de contrŰle inhibiteur diffus induit par des
stimulations nociceptives. Un systŤme inhibiteur descendant module aussi
la douleur (figure 2.7). Contrairement ŗ la thťorie du portillon, ce type
d'inhibition provient des stimulations nociceptives qui activent un systŤme
descendant mettant en jeu la substance grise pťriaquťducale, le noyau raphť
magnus et les cornes postťrieures de la moelle (14,
101). Selon ce modŤle,
une stimulation des fibres ŗ petit diamŤtre dťclenche, vers les neurones
nociceptifs des couches I et V des cornes postťrieures de la moelle, un
systŤme d'inhibition descendant issu de la substance grise pťriaquťducale
et du noyau raphť magnus. Cette activitť analgťsique est connue sous le
nom de CIDN ou ContrŰles Inhibiteurs Diffus induits par des stimulations
Nociceptives (207,208). Ces stimulations activent les neurones du bulbe
rostro-ventral qui inhibent ŗ leur tour l'activitť des neurones nociceptifs
des cornes dorsales de la moelle. Contrairement au systŤme de modulation
proposťe par la thťorie du portillon, le CIDN produit une inhibition
descendante qui ne se limite plus ŗ la rťgion stimulťe (83,
354,355).
L'application d'un stimulus nociceptif intense active le pool de neurones
nociceptifs non spťcifiques correspondant au segment mťdullaire activť,
tout en inhibant simultanťment des neurones du mÍme type situťs ailleurs
dans la moelle ťpiniŤre. En rťduisant l'activitť des neurones nociceptifs
non-spťciques, les CIDN attťnueraient le bruit de fond et feraient
ressortir l'activitť des neurones spťcifiques sollicitťs par la stimulation
nociceptive (209). Selon cette hypothŤse, la douleur ne serait pas
uniquement dťclenchťe par des processus excitateurs, mais par la perception
d'un contraste entre les activitťs de deux pools de neurones (27). Ce
modŤle explique bien l'inhibition gťnťralisťe de la douleur produite par
des stimulation intenses, voire pťnibles.
Dans le systŤme inhibiteur descendant, le blocage de la rťponse nociceptive
se produit dans trois zones diffťrentes. D'abord, dans la rťgion du
mťsencťphale oý une stimulation au niveau de la substance grise
pťriaquťducale amŤne une inhibition du message nociceptif sans influencer
la transmission des autres messages. Une stimulation de la substance grise
pťri-ventriculaire de l'hypothalamus produit ťgalement le mÍme effet (ces
deux sites sont anatomiquement reliťs). L'information parvenant au
mťsencťphale arrive principalement de la partie antťrieure du bulbe, de la
moelle ťpiniŤre et du lobe frontal. L'information provenant de la moelle
joue ici un rŰle prťpondťrant. Ensuite, l'inhibition de la rťponse
nociceptive peut se produire dans la rťgion rostro-ventrale du bulbe. Cette
rťgion comprend notamment le noyau raphť magnus et la formation rťticulťe
du bulbe. Elle reÁoit ses informations principalement de la substance grise
pťriaquťducale et de la formation rťticulťe adjacente au mťsencťphale.
Enfin, l'inhibition du message nociceptif se produit aussi dans la rťgion
latťrale et dorso-latťrale de la protubťrance annulaire. Peu ťtudiťe, cette
zone territoire envoie des projections vers la substance grise
pťriaquťducale, la partie rostro-ventrale du bulbe et la moelle ťpiniŤre.
Le funicule dorso-latťral constitue le relais descendant entre les deux
rťgions mentionnťes et la moelle ťpiniŤre. Toutefois, peu de projections
directes relient la substance grise pťriaquťducale et le funicule
dorso-latťral. La majeure partie de l'information passe probablement
d'abord par la partie rostro-ventrale du bulbe avant de rejoindre les lames
I, II et V de la moelle via le funicule dorso-latťral (100).
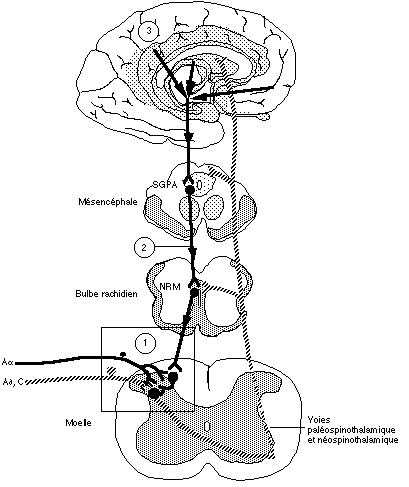
Figure 2.7 Mťcanismes de modulation de la douleur
Mťcanismes de modulation de la douleur dans le systŤme nerveux central.
1= ContrŰle mťdullaire (thťorie du portillon) un interneurone inhibiteur de
la substance gťlatineuse, recrutť par les grosses fibres non douloureuse (A-alpha),
bloque l'activitť des fibres nociceptives (A-delta et C).
2= ContrŰle inhibiteur diffus induit par des stimulations nociceptives (CIDN)
l'activation des nocicepteurs dťclenche un systŤme inhibiteur descendant
diffus. 3= ContrŰle des centres supťrieurs du systŤme nerveux central :
plusieurs structures des centres supťrieurs modulent la perception de la
douleur (SGPA: substance grise pťriaquťducale, NRM: noyau raphť magnus).
|
Deux neurotransmetteurs participent au contrŰle exercť par le systŤme
inhibiteur descendant : les transmetteurs biogťniques aminťs et les
opiacťs. Dans le cas des transmetteurs aminťs, les projections
issues de la rťgion rostro-ventrale du bulbe comprennent des neurones
sťrotoninergiques. Les zones issues de la rťgion pontine tegmentum
contiennent des neurones noradrťnergiques. Leur action mťdullaire est
rendue possible par des rťcepteurs adrťnergiques alpha2 concentrťs
dans les lames supťrieures de la moelle. ņ l'ťvidence, la sťrotonine,
issue des neurones sťrotoninergiques, agit directement sur les neurones
de la corne dorsale pour les inhiber. Pour leur part, les opiacťs figurent
parmi les analgťsiques connus les plus puissants. Leur efficacitť
s'explique en grande partie par les rťcepteurs opiacťs spťcifiques
contenus dans certaines synapses des lames supťrieures de la corne dorsale,
de la substance grise pťriaquťducale et probablement aussi de la substance
grise rostro-ventrale du bulbe, une rťgion moins bien ťtudiťe. D'un autre
cŰtť, certains neurones renferment des dťrivťs d'opiacťs libťrables lorsque
la situation le requiert. Les concentrations les plus marquťes
d'enkťphalines se trouvent dans le systŤme nerveux sympathique, l'intestin
et les cellules chromaffines des surrťnales. Les dynorphines, retrouvťes
aux mÍmes endroits mais dans des cellules diffťrentes, et les endorphines-Ŗ
en contiennent aussi.
Il reste encore ŗ dťcouvrir au niveau des mťcanismes assurant le
fonctionnement des contrŰles descendants. De nombreux neurotransmetteurs,
dont plusieurs peptides, ont ťtť identifiťs particuliŤrement au niveau de
la colonne dorsale. Certains d'entre eux y jouent probablement un rŰle
inhibiteur. Ainsi, les neurones sťrotoninergiques agissent directement et
de faÁon postsynaptique sur les cellules de la corne dorsale. Ils
influencent aussi des interneurones contenant des enkťphalines qui, en
retour, interviennent post-synaptiquement sur les cellules de la corne
dorsale. De plus, la terminaison centrale de plusieurs affťrents primaires
possŤde des rťcepteurs opiacťs. Ainsi l'interneurone contenant des
enkťphalines pourrait agir, de faÁon prťsynaptique, sur l'affťrent primaire
en limitant la dťcharge transmise ŗ la cellule de la corne dorsale.
L'application de stimuli prolongťs et intenses dťclenchent l'action
analgťsique des opiacťs. Mais le stress, la peur ou l'anxiťtť la favorisent
encore davantage !
|